ON THE PROBLEM OF NERVE PATHWAYS CROSSING
M. Kazarnovsky, Ph. D.
Abstract
Some reasonings are proposed to explain why nerve pathways cross. The leading role of vision is suggested to be the basic reason for the phenomena as considered in the process of the nervous system evolution.
Introduction
The immediate reason for this paper has been a
remark in the Dr’s. David H. Hubel’s and T.N.Wiesel’s paper
in “Scientific American” “…It is worth
remarking that no one has the remotest idea, why there should be this amazing
tendency for nervous system pathways to cross…” (Hubel DH, Wiesel TN, 1979).
In the Dr. D. Hubel’s book “Eye, Brain and Vision”,
one can find the following:
“…Incidentally,
no one knows why the right half of the world tends to project to the left half
of the cerebral hemispheres…” (Hubel DH., 1988).
The question is still unanswered. No need to explain the importance of the
problem.
The nerve pathways’ crossing is the basic structural
feature of vertebrate brain.
I
shall take a risk to put forward an idea, which in my opinion gives the
simplest explanation for the phenomena.
Methods and Materials
The leading role of sight in life evolution
The
light perception started from a primitive light sensitive spot on the skin. But,
nevertheless, this extremely important ability to distinguish light from dark
played the tremendous role in evolution. The species, which possessed this
ability, had an advantage over the ones not being able to get a light signal. Next step in vision
development has been a transformation of light sensitive spot into lens which
widened the field of vision and made an eye to be a powerful perception
mechanism leading at the final step to images forming and analyzing ability. It should be emphasized that we are talking for now
only about light perception, not forming and analyzing images. But even this
ability meant a new epoch in the evolution.
Detailed description of images perception and analyzing
is beyond the scope of the paper. One can find it in a number of sources.
Why nerve pathways cross
We
will concentrate now on the optics of the phenomena of nerve pathways crossing.
First,
I will provide a short review of the optic pathways
organization in human.
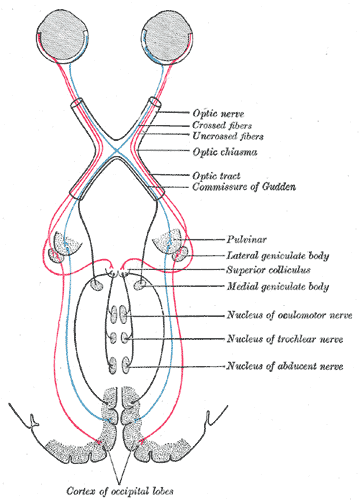
Fig.
1. Optic pathways organization
The
part of the optic pathways starting from the retinal ganglion and going up to the
optic pathways crossing point is called an “optic nerve”. The crossing point
itself is called an “optic chiasm”, and the part behind the optic chiasm up to
the cortical structures is called an “optic tract”.
At the chiasm, approximately 25%-30% (in human) of optic
nerve axons cross the chiasm to the contralateral (opposite) side of the brain where
they merge with the uncrossed axons from another eye forming an optic tract on
a contralateral side of the brain, while the rest 70%-75% of optic nerve axons
continue their way through the same side of the brain – unilaterally - as an
optic tract. As a result, after chiasm all axons representing left part of the vision field constitute
right optic tract and axons
representing right part of the vision field constitute left optic
tract.
This
means that the left part of the visual field is projected on the right part of
the cortex and visa versa.
Majority of the sensory and motion control
systems follow this contralateral organization. This is a well-known fact and I
will not go further with it.
So, the question is why the evolution chose
this contralateral way to manage a species perception-reaction control? To
approach the possible explanation let us consider a simple model, i.e., how
light penetrates a lens with sensitive layer at the bottom.
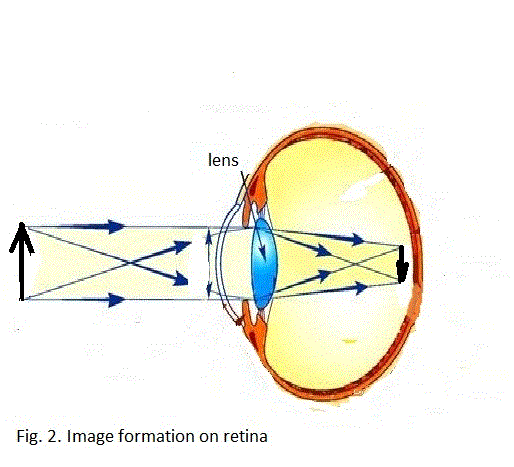
Due to the lens light refracting property,
left-right object’s configuration is inverted to the right-left configuration of
this object’s image on the sensitive layer. Note that the up-down sides are
reversed as well.
I suggest that the lens refracting property
is the basic reason for the contralateral nervous system organization.
Body structures have been developed
gradually during millions of years, vision structures included, and it could be
that the first lens leading mutation happened in one light sensitive spot first
or in a pair of them at once - it really makes no difference as both convert
the object’s image in the same way - leading to the right / left images coming from
the two eyes to merge into left / right images contralaterally.
It is logical to assume that in order to
provide a fast reaction, which can be a lifesaving, like eye and head movement
toward the moving object, it is much more efficient to start this reaction
circuit to the visual signal from the point closer to the point to where visual
information arrives, that is preferably on the same left / right side.
I would correlate this image left/right visual
reaction mechanism with the tectopulvinar system in the first place, which
remarkably works even with the damaged primary visual cortex (Ratey J, 2002),
being responsible to turn an eye in the direction of the moving object without
actually seeing it. In non-mammals the tectopulvinar system is the dominant structure for fast reaction to the visual stimulus. The
fact witnesses that the tectopulvinar system preceded geniculostriate
system and is dedicated to providing a fast reaction to the visual stimulus. That is, we can consider a tectopulvinar system as a
blue print of much more sophisticated geniculostriate system.
Accepting that the visual system has been
developed contralaterally, it is also logic to assume that another nerve structures have been developed also contra laterally to be
in sync with the vision system organization which plays the leading role in
perception. If not, there would be lots of confusion and disintegration between
different sensory systems.
I
will try to confirm this by a number of facts about sensory system
architecture.
The degree of axon’s splitting is different for the different species.
To get an idea let us take a look at the
visual system schema of a bird compared to visual system of a human being:
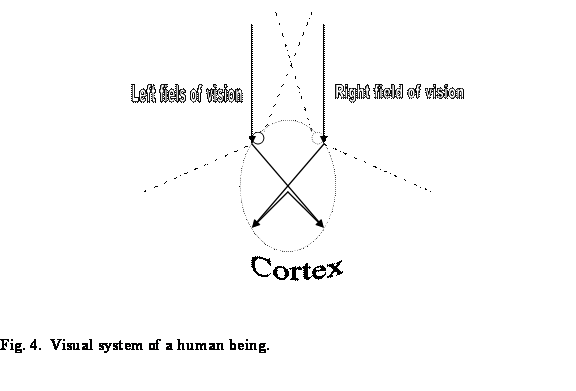

Now I assume it is not so surprising that
almost 100 percent of bird’s nerve fibers cross the chiasm and goes to the
contralateral side of the bird’s brain in contrast to human’s 25-30 percent of axons
proceeding to the opposite side of the brain and merging with the 70-75 percent
of axons originating from another eye and thus forming a full one side field of
vision. This could be explained by the obvious fact we that bird’s eyes are
located on the opposed sides of the head, which means that the two vision fields
have nothing in common and full half of the world accumulated in the cortex is
produced by the axons coming from one eye only.
“ … In birds …
optical nerve in each eye crosses over almost completely in the optic chiasm. …”. (Springer SP, Deutch G., 1997).
Another remarkable proof could be obtained from the comparison of another
sensory organs organization with the vision system organization.
Let
us take a look at human’s olfactory system for example.
“…olfactory receptor neurons, like taste buds, wind
up represented in the ipsilateral (unilateral – MK) cerebral hemisphere. Hence
the chemical senses are at the opposite end of the spectrum from somatic
sensory and motor systems, in which a given part of the body is represented
primarily in the contralateral cerebral hemisphere.” (Nolte J., 1988).
And
more: “All chemically initiated
sensory organs are unilateral” (Nolte J., 1988).
The
only reasonable explanation for that in my opinion could be that chemically
initiated sensory organs were developed before vision structures. It sounds quite reasonable as the
first primitive species had a very primitive contact with the environment via
direct physical contacts: tactile, odor, taste.
The
interesting trend could arise from here. That is, investigating the uni- contra-lateral
nerve structures organization and their relations we can trace the order in
which those structures have been developed in the process of evolution with
regard to the vision system development.
Results
Summing
up all the above, I’d like to give a support to this contra laterality idea
from the philosophical point of view, namely Ockham’s razor principle, which
states that the explanation of any phenomenon should
make as few assumptions as possible, eliminating, or "shaving off",
those that make no difference in the observable predictions of the explanatory hypothesis
or theory. This could be paraphrased as "all the things being equal,
the simplest solution tends to be the best one."
In other words,
when multiple competing theories are equal in other respects, the principle
recommends selecting the theory that introduces the fewest assumptions and
postulates the fewest hypothetical entities.
In this sense the
idea of the leading role of vision in contralateral brain structures
organization ideally fits Ockham’s razor
principle, being a simple one and at the same time not requiring any additional
assumptions.
All
kinds of critics are most welcome.
P.S. This article has been written in 2009. Recently I discovered that the same explanation
of nerve pathways crossing as a result of refracting property of the lens has
been suggested in 1899 by S. Ramon Y Cajal in the article
in German (translation from the Spanish version published earlier).
(The structure of the optic chiasm: together with a general theory of the
crossing of the nerve pathways)
I have no explanation why it was still in question in 1979 and 1988.
References
Hubel
DH,
Hubel
DH (1988) Eye, Brain and Vision, Sci.Am.Lib.
Nolte
J (1988) The Human Brain: an introduction to its functional anatomy, Mosby.
Parker
A (2003) In the blink of an eye. Perseus publishing.
Ratey
J (2002) A user’s guide to the brain. Vintage books.
Springer
SP, Deutch G.(1997) Left Brain Right Brain,
W.H.Freeman.N.Y.
Dr.
S. Ramon Y Cajal (1899) Die Structur
des Chiasma Opticum nebst einer allgemeinen theorie der kreuzung der nervenbahnen, Leipzig, Verlag von Johann Ambrosius Barth.